Percussion
Music & Motor System
Drumming, or instrumental sound generation of any
sort, is very uncommon among vertebrates. Perhaps the most striking other
example are palm cockatoos, Probosciger aterrimus, which use a stick to strike hollow
trees as a communication signal (Wood, 1984, 1988). The only other clear
examples are by woodpeckers (who sometimes drum with their bills against
particularly resonant trees as displays, in territorial/mate attractions
contexts similar to those associated with song in songbirds (Dodenhoff et al.,
2001; Stark et al., 1998). Kangaroo rats and some other desert rodents drum out
patterns with their hind feet on the ground (Randall, 1997). But bimanual
drumming per se is, as far as I know, unique to the great apes and humans.
----
W. Tecumseh Fitch (2006)
In human
cultures, singing, rapping, and dancing are
often accompanied by percussion music, which reinforces/represents the
hierarchy of the metrical structure through regular accents. Metrical structures
refer to the way musical beats are grouped, such as marches (two beats grouped together) or waltzes (three beats grouped together). The
hierarchy of the metrical structure can be regarded
as a “time ruler” with labelled graduation
marks (Figure 1). Such structures may reflect humans' superior capacity for
temporal organization.
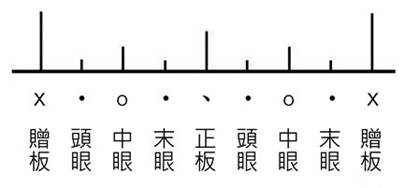
Figure 1. A “time ruler” with labelled graduation
marks and the metrical symbols in Kunqu.
We conducted
two brain-imaging experiments on the rhythmic patterns of percussion music
(Tsai et al., 2010; 2012). In the first experiment, we compared the brain
activation patterns associated with the learned percussion music of Beijing
opera and the unlearned percussion music of Sichuan opera. The participants
recruited the premotor cortices (PMC) and the pre-supplementary motor area
(pre-SMA) to covertly recite along with the learned percussion music (Figure
2). In the second experiment, we examined the brain activation patterns
associated with the rhythmic patterns of the drum set in rock music. The
participants (skilled drummers) did not recruit the pre-supplementary motor
area to covertly recite along with the learned percussion music. Given that the
percussion music of Beijing opera is characterized by subtle timing
manipulations and shifts between metrical structures, we suggest that the
pre-SMA plays a key role in error-monitoring during subvocal rehearsals. This
view is consistent with previous experiments demonstrating error-related
activity in the SMA/pre-SMA during motor tasks (Gallea et al., 2008; Klein et
al., 2007). More investigations are needed to compare different genres and
traditions of human percussion music.
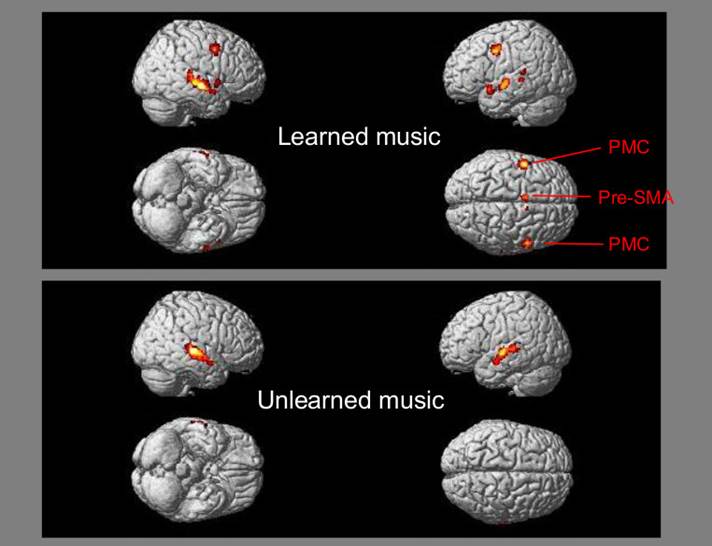
Figure 2. A comparison of
the brain activation patterns associated with the learned percussion music of
Beijing opera and the unlearned percussion music of Sichuan opera (Tsai et al.,
2010).
REFERENCES
Dodenhoff, D.
J., Stark, R. D., & Johnson, E. V. (2001). Do woodpecker drums encode
information for species recognition? Condor, 103, 143–150.
Fitch, W. T.
(2006). The biology and evolution of music: a comparative perspective.
Cognition, 100, 173–215.
Gallea, C.,
Graaf, J. B., Pailhous, J., & Bonnard, M. (2008). Error processing during
online motor control depends on the response accuracy. Behavioural Brain
Research, 193, 117–125.
Klein, T. A.,
Endrass, T., Kathmann, N., Neumann, J., von Cramon, D. Y., & Ullsperger, M.
(2007). Neural correlates of error awareness. Neuroimage, 34, 1774–1781.
Randall, J. A.
(1997). Species-specific footdrumming in kangaroo rats: Dipodomys ingens, D.
deserti, D. spectabilis. Animal Behaviour, 54, 1167–1175.
Stark, R. D.,
Dodenhoff, D. J., & Johnson, E. V. (1998). A quantitative analysis of
woodpecker drumming. Condor, 100, 350–356.
Tsai, C. G.,
Chen, C. C., Chou, T. L., & Chen, J. H. (2010). Neural mechanisms involved
in the oral representation of percussion music: an fMRI study. Brain and
Cognition, 74, 123-131.
Tsai, C. G.,
Fan, L. Y., Lee, S. H., Chen, J. H., & Chou, T. L. (2012). Specialization
of the posterior temporal lobes for audio-motor processing - evidence from a
functional magnetic resonance imaging study of skilled drummers. European
Journal of Neuroscience, 35, 634–643.
Wood, G. A.
(1984). Tool use by the Palm Cockatoo Probosciger aterrimus during display.
Corella, 8, 94–95.
Wood, G. A.
(1988). Further field observations of the Palm Cockatoo Probosciger aterrimus
in the Cape York Peninsula, Queensland. Corella, 12, 48–52.
[Home]