
![]()
|
Assessing
the Effect of Spotted Tentiform Leafminer Injury to Apples- An
oral presentation for the Centennial
Conference of the American Society for Horticultural Science October
2003, Providence, Rhode Island, USA(2.8MB) Carbon
Balance Dynamics to Intergrate Whole Tree Physiology and Cultural Practices-
An oral presentation for the Department of Horticulture at National
Taiwan University May 27, 2002 (4MB) Physiological
Effect of Summer Pruning in Apple Trees-An oral presentation
for Great Lakes Fruit Workers Conference November 30, 2000, Ithaca,
New York, USA (250KB) Summer
Pruning and Re-exposure Effects on Leaf and Whole Canopy Gas Exchange
in Apple Trees-An oral presentation for the 7th International
Symposium on Orchard and Plantation Systems February 2000, Nelson, New
Zealand (presented by Dr. Alan N. Lakso) Summer
Pruning Effects on Leaf and Whole Canopy Gas Exchange in Apple Trees-An
oral presentation for the 96th International Conference of the American
Society for Horticultural Science June 1999, Minneapolis, Minnesota,
USA Summer
Pruning Effects on Canopy, Root System, and Integral Carbohydrate Balance
in Apple Trees-A
grant proposal submitted to the Arthur Boller Apple Research Grants
1999 |

|
Summer Pruning Effects on Canopy, Root System, and Integral Carbohydrate Balance in Apple Trees
Summer pruning is commonly used in the modern apple culture system in New York State to control tree size and increase the light penetration into inner canopy to improve fruit color. However, summer pruning may reduce fruit size, yield and the consequently commercial profit. This effect is probably due to the decrease in carbohydrate supply after sizable quantities of leaf area are removed by summer pruning. Our prior studies showed that a significant decrease in whole-canopy net carbon dioxide exchange rate can be induced by a commercial pruning severity. However, the inconsistent influence on fruit growth in our studies suggested that the reproductive growth did not response immediately to the decrease in carbon supply. Based on the concept of carbohydrate balance we hypothesize that other carbohydrate sinks such as root system, trunk, and carbon reserves may be more sensitive to summer pruning but may require successive years of summer pruning to reduce tree vigor and productivity. Therefore, it is necessary to exam many components of carbohydrate supply/demand to get a integrated overview of summer pruning effects. This proposed research will examine the influences of summer pruning on root system growth, canopy photosynthesis, and whole-plant respiration to estimate the short and long term effects of summer pruning . Our goal is to develop a simulation model to integrate and quantify the effects of this pruning practice into the production system to maintain sustainability and profitability of commercial apple management.
The
objectives of this proposed research are:
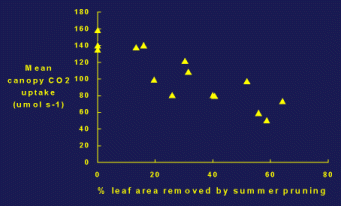
Summer pruning has become a routine practice in modern apple orchard management to improve fruit color and control tree size. This is especially important for varieties grown where the weather is not ideal for color development, such as New York State (Fig.1). Because exposure to sun light is important for fruit color development, by removing extension shoots from the outer canopy, summer pruning increases light penetration into the canopy and to the fruit, hence increases the fruit color and percentage of well-colored fruit number. On the other hand, the removal of extension shoots with large vigorous and well exposed leaves is expected to reduce the overall carbon dioxide assimilation by the tree. This would be a disadvantage for fruit growth if the carbohydrate demand is high, i.e. when the crop load is high, resulting in smaller fruit size or yield reduction. Our previous studies in 1998 have shown that the re-exposure of shading leaves after summer pruning did not functionally compensate for the reduction in carbohydrate supply lost from the removal of extension shoots and leaves. The decreases in whole-canopy net carbon exchange rate (NCER) were approximately proportional to pruning severity and percentage leaf removal (Fig. 2). However, data from our studies show the influence of summer pruning on fruit growth in the same year is somewhat ambiguous and did not fully reflect the decrease in NCER. Based on the carbohydrate balance concept, this result suggests that not only fruit but other carbohydrate sinks should also be affected by the reduction in carbon supply after summer pruning. Therefore, we hypothesize that the removal of extension shoots and leaves by summer pruning reduces the whole-tree respiration, carbon reserves, trunk and root system growth. In consequence, although summer pruning may not affect fruit growth in the same growing season, its influence on vegetative growth and flower bud development are probably additive after successive summer pruning practices. Summer pruning that reduces canopy NCER will likely become significant on next year or long-term tree vigor and productivity. Maintaining a constant annual cropping and stable fruit quality is a important issue for sustainable management of a modern commercial orchard. By analyzing each component in the carbohydrate supply/demand the proposal will provide information for integrating the influences of summer pruning and optimizing this practice.
Literature
Review
Material and pre-summer pruning easurements-twenty-year-old, slender spindle 'Empire' trees with similar bloom density and vigor will be selected in May, 1999. Crop load of each tree will be hand-regulated after fruit set to maintain a final target range. Twenty fruits at different canopy location on each tree will be labeled for monitoring fruit growth. Total leaf area of each tree will be estimated when the canopies are fully expanded before summer pruning. Four clear root observation tubes will be mounted around the rhizosphere of each tree before new roots start growing in the early season for observing root growth (Fig. 3; Caldwell and Virginia, 1991; McMichael and Taylor, 1987). In early August right before summer pruning is applied, light interception of each tree will be measured by a ceptometer line light sensor. Fish-eye photography will be used for analyzing light penetration into different canopy locations. Whole-canopy photosynthesis and dark respiration will be measured by whole-canopy balloons and a automated gas exchange system (Fig. 4; Lakso et al., 1999). Root growth will be monitored by a video periscopy. Root/soil respiration will be measured by inverted chambers and gas exchange system.
Pruning treatments-Trees will be randomly assigned to different pruning severity treatments in early August when the extension shoots reach maximum length and the leaves are fully expanded. The pruning severity is determined by the proportion of extension shoots thinned out from the canopy. Removed shoots and leaves from each tree will be collected to measure the percentage of total leaf area reduced by summer pruning Post-summer pruning measurements-Measurements mentioned earlier will be repeated right after summer pruning and later in the growing season to monitor the change and response of each component to the pruning treatment. Fruit quality analysis-Fruits will be harvested on early or October for fruit number and total yield. Twenty fruits will be randomly sampled right after harvest for quality analysis including fruit weight, color, firmness, starch and soluble solids content. Twenty other fruits will be cold stored at least 4 months to exam the internal browning and the other storage characteristics. Return cropping and root growth-The number of flower clusters and final cropping on each tree will be recorded in May to June, 2000. Re-growth of the root system will also be measured in the early growing season. Development of a simulation model-Information collected from this year and previous experiments will be integrated into the simplified dry matter production model developed by Lakso and Johnson (1990) with "Stella" dynamic simulation automatic programming language to exam the feasibility of quantifying and predicting summer pruning effects and with the simulation model.
|



|
|
|
Brought
you by:
|