![]()
![]()
![]()
![]()
![]()
![]()
我 們實驗室的研究興趣在於植物演化中,有關系統分類、分子演化、植物生殖系統的演化等研究,同時也進行一些跨領域研究的課題。近年來的研究主軸包含了寄生植 物的演化,植物花瓣/類花瓣的演化發生學研究,以及植物在生殖策略上的性別表現與器官功能轉換(transference of function)等,以下是研究室幾個主要的研究方向:
 屋久島蛇菰(大圖)和日本蛇菰(小圖)都只具有雌性個體  帽蕊草的雄蕊特化成筒狀,蓋在雌蕊上,成熟後會整個掉落,露出柱頭(如圖所示) |
(1) 寄生植物之系統分類與演化:寄生植物在被子植物中平行演化超過十次以上,包含了全寄生和半寄生的各式物種,以全寄生植物而言,除了因不會行光合作用而沒有綠色組織 之 外,在花部的形態上也多半相當特異,因而在分類鑑定上有許多困難之處。我們實驗室目前特別針對蛇菰科(Balanophoraceae)和帽蕊草科 (Mitrastemonaceae)在進行研究,包含對於其系統分類,生殖生物學,以及基因組演化等課題。 蛇菰科的植物約有17屬120種,其中最大的屬是蛇菰屬(Balanophora), 目前認定的有16-17種,但是有近百個學名曾被發表。蛇菰屬的分布自非洲東部,東亞,東南亞,一直到澳洲北部;但是物種多樣性集中在東亞到東南亞這個區 域。臺灣目前已知的種類包含疏花蛇菰(B. laxiflora), 筆頭蛇菰(B. harlandii),海桐生蛇菰(B. tobiracola),粗穗蛇菰(B. fungosa ssp. fungosa),和最近確認的日本蛇菰(B. japonica)與屋久島蛇菰(B. yakushimensis)等六種(Su et al. 2012)。蛇菰屬植物還有一個特性即是基因組呈現高度變異的情形,以rDNA序列而言,其核酸取代速率較其他被子植物高出6倍以上(Su & Hu 2012)。因此針對其核和胞器基因組的演化有許多有趣的研究課題。 帽蕊草科植物只有帽蕊草屬(Mitrastemon), 全世界只有2-3種,分布在太平洋的兩邊。帽蕊草(Mitrastemon yamamotoi)複合群分布在東亞到大洋洲,而松田氏帽蕊草則分布在中南美洲。前者在臺灣多半鑑定成兩種:臺灣奴草(M. kawasasakii和菱形奴草(M. kanehirai), 但可供區分的形態以植株大小為主,而常有中間型態產生,並不穩定。帽蕊草在過去曾被許多分類學者放在大花草科(Rafflesiaceae)之中,在近年 的APG III系統中,則確定應該自成一科,並位在杜鵑花目(Ericales)中,而與黃褥花目(Malpighiales)中的大花草科親緣關係不大。我們目 前希望分析其基因組,以確定其質體(plastid)的存在,並瞭解它在杜鵑花目中的確切分類位置。
|
 蕘花葉灰木的「兩性花」中的五枚雄蕊都是「空包彈」,沒有花粉的產生。 |
(2) 植物生殖系統之演化:以被子植物來說,花多半是兩性的(bisexual, hermaphroditic),但是單性花在植物界中也非常常見。單性花包含了雌雄同株(monoecy)和雌雄異株(dioecy)兩大類,後者即和 動物系統中的雌雄性相類似。但是雌雄異株植物則只佔了整體被子植物的4-6%而已,並且在植物演化中平行演化多次。然而過去的研究(如Bawa等人)發現 到一個有趣的現象,就是熱帶島嶼的雌雄異株植物比例比其他地區都來得高。我們研究室針對台灣的植物也做了一些觀察分析,發現臺灣植物中,雌雄異株的比例大 概在7.9%左右,而其中亞熱帶雨林區域更高達24%(Tseng et al. 2008)。上述的疏花蛇菰、寶島玉葉金花等都是雌雄異株的種類。 然而對於臺灣植物的性別系統表現我們知道的依然十分有限,比方說灰木科的蕘花葉灰木(Symplocos wikstroemia ) 在外形中有開雄花和雙性花的不同植株。但在進一步的研究中,我們發現它其實是隱性的雌雄異株植物,其「雙性花」只有雌性的功能,它的花藥並沒有花粉的發育 (Wang & Hu 2011)。有關其他可能的隱性雌雄異株物種,或是被稱為具有「雜性花(polygamic flowers)」物種的研究,則有待更多的投入。 另外有關被子植物中的孤雌生殖(agamospermy)現象也較少被提及或重視。先前所提過的日本蛇菰和屋久島蛇菰都被認為是孤雌生 殖的種類,而我們也發現在臺灣的蕁麻科樓梯草屬(Elatostema) 中,也有三種是孤雌生殖的物種。 |
 硃砂根的葉子邊緣有瘤狀腫大,即為有共生菌生長的葉瘤 |
(3) 紫金牛屬植物與其共生細菌的演化關係:植物與微生物的共生關係在過去的研究多集中在豆科植物與固氮根瘤菌中,然而此種關係需每代建立,其共譜系關係往往不明顯。目前已發現有 些植物可以與細菌共生形成共生葉瘤,並將細菌包入種子中,成為循環式共生。此現象目前只被發現在部份茜草科、紫金牛科和薯蕷科植物中。在紫金牛屬圓齒紫金 牛亞屬(Crispardisia)的例子裡,細菌存在於葉緣的葉 瘤以及頂芽基部黏液中,在開花結果後隨著種子傳到下一代,故成為一個特殊的共生關係。我們實驗室先前已成功利用PCR鑑定台灣紫金牛屬植物之內共生細菌, 並初步證實共譜系關係的可能存在(Ku & Hu 2014)。目前則希望擴大研究至其他圓齒紫金牛亞屬的成員,以及瞭解植物與細菌的實際互動情形與機制。 |
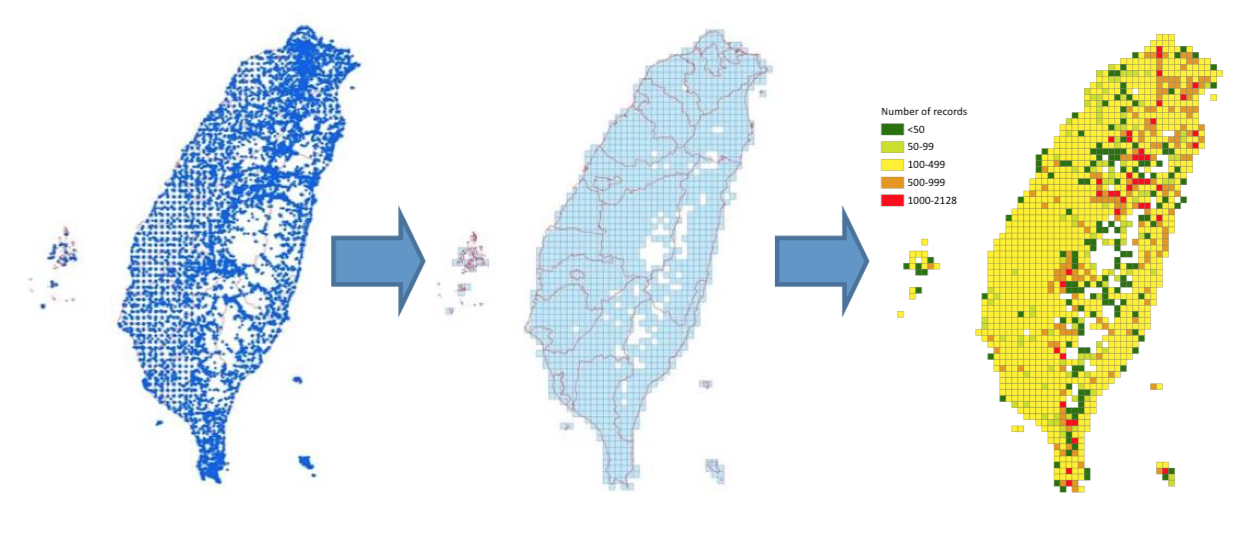 物種分布資料 |
(4) 台灣植物分布類型研究:植物分布的類型與環境本身生物或非生物的因子,以及生物歷史的脈絡都有關係。我們希望利用大量相關的數據,建構出台灣植物分布類型,並對特有性,稀有性等進行研究。整個資料庫的建構包含各大標本館的標本資料,以及數次植物調查的樣區資料等。希望呈現台灣植物的分布特性。 |
 阿美族利用許多不同的野菜 |
(5) 民族植物研究:民族植物與傳統生態智慧的相關研究在臺灣近年漸漸受到許多重視,然而對於全球化與文明化的衝擊,許多自然環境 與經年累月所孕積的生態智慧都不斷的被破壞消磨,甚至逐漸被遺忘。我們研究室與台大人類系的羅素玫老師目前合作進行台東都蘭阿美族的野菜園研究,希望以此 做為更多人投入民族植物研究的契機。 |
 台大植物標本館(TAI herbarium) |
(6) 臺灣的植物探索史研究:臺灣自十九世紀以來,經歷不同時期的殖民統治,也有來自歐洲與日本的諸多生物學者在台灣進行採集與植物研究。 我們研究室對於日治時期,特別是臺北帝大時期(1928-1945)的植物研究特別有興趣,目前正針對臺北帝大時期太平洋植物資源調查,配合植物標本 館的館藏進行整理,希望重現這部份史料與標本的真實面貌。日治時期早期,由川上瀧彌、金平亮三,及東京帝大的早田文藏等學者的努力下,奠定了堅實的基礎。台北帝大時期的植物分類研究學者,由原北海道大學的工藤祐 舜教授領軍,開創了臺北帝大的植物學講座,然而在工藤教授的英年早逝的影響下,植物分類研究發展一度受到重創,而在正宗嚴敬、山本由松、細川隆英、鈴木重 良等人的努力下,植物學的研究終於走出臺灣自己的天空,甚至觸腳伸到東南亞和太平洋島嶼。 |
 昆欄樹的花沒有明顯的花被構造  寶島玉葉金花(Mussaenda formosana)具有白色 的膨大萼片。 |
(7) 植物花被的演化研究:植物為確保有性生殖的遂行,有著豐富多變的花朵構造 來幫助傳粉作用。有些植物沒有明顯花被,而有些植物並非利用單一朵花,而是利用整個花序形成假單花(pseudanthium)來吸引傳粉者,比如大部份 的菊科植物,利用頭狀花序來模擬單一朵花。 根據模式植物阿拉伯芥和金魚草等所建立的花部發育ABCDE模型顯示,控制花瓣的器官形成是由ABE三群基因共同來決定的因此,研究這些基因應有助於瞭解 花被的演化。 我們研究室選擇了一些花被有特異演化的植物進行比較研究,一是不具明顯花被的昆欄樹(Trochodendron aralioides),還有黃楊科的黃楊(Buxus microphylla),以及花被僅存於雄花而雌花缺如的蛇菰科蛇菰屬 (Balanophora spp.)植物。我們首先確定ABCDE群基因在這些植物中的存在,並觀察它們的表現型式。昆欄樹的相關研究成果已發表於International Journal of Plant Sciences (Wu et al. 2007)。研究結果顯示昆欄樹雖無明顯花被,但ABCDE群基因都仍功能完好,然而其表現形式和阿拉伯芥等相比還是有些許差異。我們也和哈佛大學的 Elena Kramer實驗室共同合作有關MADS-box基因的分子演化(Kramer et al. 2006)。 另一方面,我們選擇了一些具有假單花構造且為類花瓣(petaloid)的植物,意即利用非花瓣的器官來模 擬花瓣的植物,例如山茱萸科四照花(Cornus kousa)的膨大苞片、茜草科寶島玉葉金花(Mussaenda formosana和八仙花科的華八仙(Hydrangea chinensis)的膨大花萼等。由於類花瓣在被子植物的演化歷史 上獨立演化了許多次,因此如何在許多不同的例子中尋求功能轉換的一致性便是一個有趣而重要的研究課題。 |
其他:
「科學教育」:大學博物館行動展示盒的研發
References
Ku, C. and J.-M. Hu*. 2014. Phylogenetic and cophylogenetic analyses
of the leaf-nodule symbiosis in Ardisia
subgenus Crispardisia
(Myrsinaceae): evidence from nuclear and chloroplast markers and
bacterial rrn operons. International Journal of Plant Sciences 175(1):
92-109.
Kramer, E. M.*, H.-J. Su, C.-C. Wu, and J.-M. Hu. 2006. A simplified explanation for the frameshift mutation that created a novel C-terminal motif in the APETALA3 gene lineage. BMC Evolutionary Biology 6: 30.
Su, H.-J. and J.-M. Hu*. 2012. Rate heterogeneity in six protein-coding genes from the holoparasite Balanophora (Balanophoraceae) and other Santalales taxa. Annals of Botany 110: 1137-1147.
Su, H.-J., J. Murata, and J.-M. Hu*. 2012. Morphology and
phylogenetics of two holoparasitic plants, Balanophora japonica and Balanophora yakushimensis
(Balanophoraceae), and their hosts in Taiwan and Japan. Journal of
Plant Research 125: 317-326.
Tseng, Y.-H., C.-F. Hsieh, and J.-M. Hu*. 2008. Incidences and
ecological correlates of dioecious angiosperms in Taiwan and its
outlying Orchid Island. Botanical Studies 49: 261-276.
Wang, Y.-C. and J.-M. Hu*. 2011. Cryptic dioecy of Symplocos wikstroemiifolia Hayata
(Symplocaceae) in Taiwan. Botanical Studies 52: 479-491.
Wu, H.-C., H.-J. Su, and J.-M. Hu*. 2007. The identification of A-, B-, C-, and E-class MADS-box genes and implications for perianth evolution in the basal eudicot Trochodendron aralioides (Trochodendraceae). International Journal of Plant Sciences 168:775-799.
[EMAIL: jmhu@ntu.edu.tw]